

User-Friendly Approach to Gain Isolation of Interphase Cells for Fluorescence in Situ Hybridization
Thomas Liehr * ![]() , Monika Ziegler
, Monika Ziegler ![]() , Nadezda Kosyakova
, Nadezda Kosyakova ![]() , Ahmed B.H. Al-Rikabi
, Ahmed B.H. Al-Rikabi ![]()
Jena University Hospital, Friedrich Schiller University, Institute of Human Genetics, Am Klinikum 1, D-07747 Jena, Germany
* Correspondence: Thomas Liehr![]()
Received: March 16, 2018 | Accepted: April 25, 2018 | Published: May 18, 2018
OBM Genetics 2018, Volume 2, Issue 2 doi:10.21926/obm.genet.1802021
Academic Editors: Joep Geraedts
Special Issue: Applications of Fluorescence in Situ Hybridization
Recommended citation: Liehr T, Ziegler M, Kosyakova N, Al-Rikabi ABH. User-Friendly Approach to Gain Isolation of Interphase Cells for Fluorescence in Situ Hybridization. OBM Genetics 2018;2(2):021; doi:10.21926/obm.genet.1802021.
© 2018 by the authors. This is an open access article distributed under the conditions of the Creative Commons by Attribution License, which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly cited.
Abstract
Background: Fluorescence in situ hybridization (FISH) can be performed on metaphase- and/or interphase cells. Metaphase cells can exclusively be obtained and studied after time-consuming cell culture. Therefore, this technique is suitable only for cells derived from peripheral blood, bone marrow or fibroblasts, the latter including skin, tumour or “prenatal tissues”, such as the amnion or chorion. On the other hand, interphase cells can be gained from many different cells and even without cell culture.
Methods: Here, we report an approach by which interphase cells can be easily obtained from the skin surface, simply and without causing any pain or injury to the patient. This straightforward procedure involves adding a few microliters of fluid to the patient’s skin (e.g. the forearm) and subsequently, using the edge of a slide to collect skin cells together with the fluid by carefully scratching along the wetted skin region. The collected fluid is distributed across the slide surface and air-dried.
Results: Applying this simple cell collection approach, several hundred skin cells are obtained. The result can be evaluated by phase contrast microscopy and regions with the highest density of cells can be selected for interphase-FISH. Here, we provide proof-of-principle of this technique. Also, the cells obtained using the described method can be used for DNA-based molecular genetic tests.
Conclusions: A new and straightforward method for acquisition of interphase cells is presented. It can be used to complement the available approaches for the analysis of different tissues in cases of questionable low-level mosaics detected in peripheral blood testing.
Keywords
Skin; interphase; fluorescence in situ hybridization (FISH); non-invasive procedure
1. Introduction
Mosaicism can be observed in all humans, particularly among females since they represent a mosaic of cells in which either the maternal or paternal X-chromosome is inactivated [1]. It has also been shown that the copy number variants present in different cell populations increases with the age of the individual [2]. Furthermore, functional mosaics due to mono-allelic gene expression seem to be the rule rather than the exception [3]. In addition to this type of physiological mosaicism, this phenomenon is also observed in relation to human pathology, especially at the chromosomal level. Examples are cases of (partial) trisomy, such as Down syndrome, Klinefelter syndrome, Triple-X syndrome, or cases with small supernumerary marker chromosomes [4,5], as well as cases with monosomy X or mosaicism in microdeletion syndromes [6,7], including tissue-specific mosaicism [8,9,10,11].
Although mosaic patterns may vary between different tissue types in a single patient [12], cytogenetic analysis is routinely performed on only one tissue and in most cases, from peripheral blood, while analysis of fibroblasts is rare. Examples can be found easily in the literature, where only few or even no aberrant cells were detected in lymphocytes, despite detection of a high percentage of such cells in skin fibroblasts.
Although a few methods for the acquisition of fibroblasts for subsequent cell culture are available, these involve invasive and painful procedures [13,14]. Therefore, we have developed a straightforward, non-invasive and pain-free protocol for obtaining cells from a patient’s skin that can be used for targeted interphase fluorescence in situ hybridization (FISH) and even for DNA-based tests. Thus, suspected small cell mosaics detectable by molecular cytogenetics can be verified in a second tissue by using this approach.
2. Materials and Methods
2.1 Cell Acquisition
In this study, we investigated three subjects; two healthy males and one healthy female. Cells obtained from all three probands were analyzed by FISH and molecular genetics as described in 2.2 and 2.3. Each experiment was repeated twice. When a male was tested, the probe was acquired by a female and vice versa.
For obtaining cells, approximately 200 of µl clean water were placed on a region of skin that was largely free of hair; the underside of the forearm is ideal and should be stretched out and presented horizontally by the proband. After adding the fluid, the long edge of a slide was used to abrade skin cells superficially. The cells and fluid were collected by carefully moving the sharp edge of the slide along the skin as shown in Figure 1A. It is recommended that this should be performed first on the experimenter’s own skin to get a feeling for the movements before applying the technique to the subject. Subsequently, the slide was brought into a horizontal position and the drop of fluid was spread with a second clean slide by the same method as that used for a blood smear. The slide was then dried in the horizontal position at room temperature and used directly for FISH. At this stage, it would also be possible to send the slide to the laboratory performing this analysis.
2.2 Fluorescence in Situ Hybridization (FISH)
FISH was performed according to standard procedures [15]. In this study, three commercially available probes (Abbott/Vysis, Wiesbaden, Germany) were applied in a three color-FISH-experiment; i.e. probes for chromosomes X (DXZ1 on Xp11.1~q11.1, green), Y (DYZ3 on Yq12, red) and 10 (D10Z1 on 10p11.1~q11.1, blue).
2.3 Y-chromosome Specific PCR Polymerase Chain Reaction
For proof-of-principle and to confirm the absence of probe contamination by the person collecting the fluid as described in 2.1, the following test was also performed. The fluid with cells collected on the slide (see 2.1.) was not spread and air-dried on the slide surface, but instead, was collected with a micropipette and used for polymerase chain reaction (PCR) analysis.
The collected fluid was concentrated to approximately 1 µl using a centrifugal evaporator, and the DNA from the sample was amplified by degenerate-oligonucleotide-primed PCR (DOP-PCR) [16]. Subsequently, 1 µl of the DOP-PCR product was used in a Y-chromosome specific PCR-reaction according to a previously described method [17].Three of the authors of this paper (TL, ABHA; NK) acted as volunteer participants in this study.
3. Results
The approach reported here is suitable for collection of minimal numbers of skin cells without causing any pain or damage to the skin. Several hundred cells and their DNA can be obtained for single cell analysis by FISH (Figure. 1B. The results were confirmed by DOP-PCR amplification (data not shown) since the amount of DNA obtained from such a few cells are too small for subsequent direct PCR-based analysis. After this step, it is possible to perform a standard PCR amplifying short DNA fragments, such as the 170-kb fragment derived from the Y-chromosome amplified in this study (Figure. 1C).
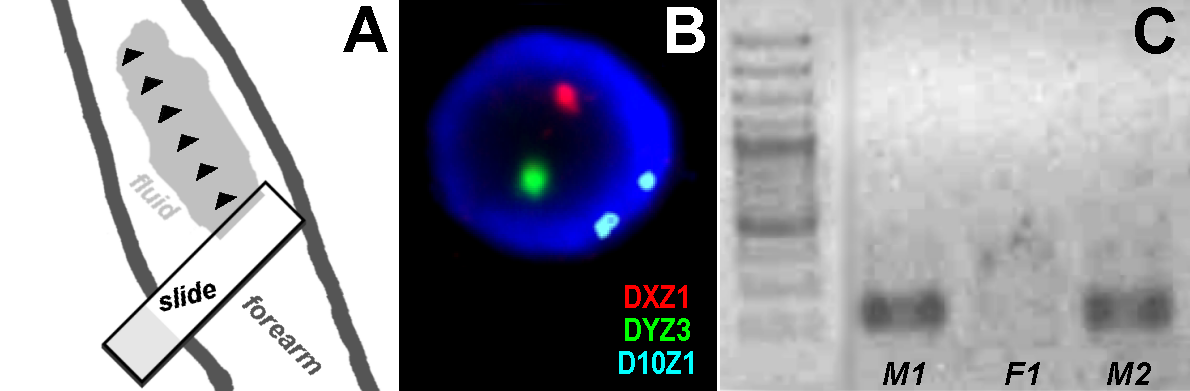
Figure 1 (A) Schematic diagram of the procedure used for skin abrasion using the long edge of a slide without causing damage; the black arrowheads indicate the direction that the slide should be moved carefully along the skin surface. (B) FISH result from male 1 using three color FISH with centromeric probes for chromosomes X (DXZ1) and 10 (D10Z1) and a probe for Yq12 (DYZ3). (C) Results of Y-chromosome specific PCR-reaction in male 1 (M1), male 2 (M2) and female 1 (F1). Clearly visible Y-chromosome material was detectable only in the two male probands; first lane: 150-bp ladder.
4. Discussion
Interphase-FISH is an important tool used in human cytogenetic diagnostics. This technique plays a major role in follow-up studies of hematological malignancies, in pathology or in prenatal diagnostics [14,18]. Research based interphase-FISH is also widely used [14,18,19,20,21].
Some examples of cases in which different tissues are necessary for clarifying clinical diagnostics are listed as follows:
-
Minimal or no clinical signs, but with a high rate of cells with an additional chromosome copy in peripheral blood [4,14,22];
-
Major symptoms but with no, or only a few, cells with an additional chromosome in peripheral blood [13];
-
No major symptoms but with diagnosed infertility together with low-level mosaicism of a numerical gonosomal aberration [6].
The new approach described here may be helpful in all these situations.
5. Conclusions
The protocol described here (previously mentioned only briefly by Liehr [15] provides an additional approach to the study of cell types other than peripheral blood in patients with a suspected or confirmed mosaic karyotype. To date, protocols for this purpose were available only for analysis of buccal smears, hair root cells or urine [15]. Thus, the protocol described here is an important additional tool that can be implemented to study different tissues in patients with molecular cytogenetically detectable conditions.
Author Contributions
TL provided the idea and developed the experiments as well as wrote the paper. MZ and ABHA performed the FISH-experiments. NK performed the molecular genetic experiments.
Competing Interests
The authors have declared that no competing interests exist.
References
- Rinčić M, Iourov IY, Liehr T. Thoughts about SLC16A2, TSIX and XIST gene like sites in the human genome and a potential role in cellular chromosome counting. Mol Cytogenet. 2016; 9: 56. [CrossRef]
- Mkrtchyan H, Gross M, Hinreiner S, Polytiko A, Manvelyan M, Mrasek K, et al. The human genome puzzle - the role of copy number variation in somatic mosaicism. Curr Genomics. 2010; 11: 426-431. [CrossRef]
- Zakharova IS, Shevchenko AI, Zakian SM. Monoallelic gene expression in mammals. Chromosoma. 2009; 118: 279-290. [CrossRef]
- Bernard R, Giraud F. [CHROMOSOME MOSAICS]. Marseille Med. 1964;101:601.
- Liehr T, Karamysheva T, Merkas M, Brecevic L, Hamid AB, Ewers E, et al. Somatic Mosaicism in Cases with Small Supernumerary Marker Chromosomes. Curr Genomics. 2010; 11: 432-439. [CrossRef]
- Liehr T, Mrasek K, Hinreiner S, Reich D, Ewers E, Bartels I, et al. Small Supernumerary Marker Chromosomes (sSMC) in Patients with a 45,X/46,X,+mar Karyotype – 17 New Cases and a Review of the Literature. Sex Dev. 2007; 1: 353-362. [CrossRef]
- Liehr T, Schreyer I, Kuechler A, Manolakos E, Singer S, Dufke A, et al. Parental origin of deletions and duplications - about the necessity to check for cryptic inversions. Mol Cytogenet. 2018; 11: 20. [CrossRef]
- Iourov IY, Vorsanova SG, Yurov YB. Chromosomal variation in mammalian neuronal cells: known facts and attractive hypotheses. International Review of Cytology. 2006; 249: 143-191. [CrossRef]
- Iourov IY, Vorsanova SG, Yurov YB. Somatic Genome Variations in Health and Disease. Curr Genomics. 2010; 11: 387-396. [CrossRef]
- De S. Somatic mosaicism in healthy human tissues. Trends Genet. 2011;27(6):217-223. [CrossRef]
- Dumanski JP, Piotrowski A. Structural genetic variation in the context of somatic mosaicism. Methods Mol Biol. 2012; 838: 249-272. [CrossRef]
- Fickelscher I, Starke H, Schulze E, Ernst G, Kosyakova N, Mkrtchyan H, et al. A further case with a small supernumerary marker chromosome (sSMC) derived from chromosome 1--evidence for high variability in mosaicism in different tissues of sSMC carriers. Prenatal Diag. 2007; 27: 783. [CrossRef]
- Liehr T, Wegner RD, Stumm M, Joksić G, Polityko A, Kosyakova N, et al. Pallister-Killian Syndrome: Rare Phenotypic Features and Variable Karyotypes. Balk J Med Genet. 2008; 11: 65-67. [CrossRef]
- Liehr T. Small supernumerary marker chromosomes. [Available from: http://ssmc-tl.com/sSMC.html] [accessed 16/03/2018]
- Liehr T. Fluorescence in situ Hybridization (FISH) – Application Guide. 2nd Ed. ed. Berlin: Springer; 2017.
- Liehr T, Weise A, Heller A, Starke H, Mrasek K, Kuechler A, et al. Multicolor chromosome banding (MCB) with YAC/BAC-based probes and region-specific microdissection DNA libraries. Cytogenet Genome Res. 2003; 97: 43-50. [CrossRef]
- Choi SK, Kim JW, Park SY, Kim YM, Kim JM, Ryu HM, et al. Retroactive DNA analysis for sex determination and dystrophin gene by polymerase chain reaction with archived cytogenetic slides. Exp Mol Med. 1999; 31: 36. [CrossRef]
- Yurov YB, Vorsanova SG, Iourov IY. Human interphase chromosomes: biomedical aspects. 2013.
- Rouquette J, Cremer C, Cremer T, Fakan S. Functional nuclear architecture studied by microscopy: present and future. Int Rev Cell Mol Biol. 2010;282:1-90. [CrossRef]
- Manvelyan M, Hunstig F, Bhatt S, Mrasek K, Pellestor F, Weise A, et al. Chromosome distribution in human sperm – a 3D multicolor banding-study. Mol Cytogenet. 2008; 1 : 25. [CrossRef]
- Manvelyan M, Kempf P, Weise A, Mrasek K, Heller A, Lier A, et al. Preferred co-localization of chromosome 8 and 21 in myeloid bone marrow cells detected by three dimensional molecular cytogenetics. Int J Mol Med. 2009; 24: 335-341.
- Papoulidis I, Kontodiou M, Tzimina M, Saitis I, Hamid AB, Klein E, et al. Tetrasomy 9p mosaicism associated with a normal phenotype in two cases. Cytogenet Genome Res. 2012; 136: 237-241. [CrossRef]
