

The Induction of Histone H3K4 Methylation on the SI Gene Correlates with SI mRNA Levels in Enterocyte-Like Caco-2 Cells
Kazuki Mochizuki 1, 2,*![]() , Ayumi Osaki 2
, Ayumi Osaki 2![]() , Yuko Inamochi 2
, Yuko Inamochi 2![]() , Toshinao Goda 2
, Toshinao Goda 2![]()
- Faculty of Life and Environmental Sciences, University of Yamanashi, Yamanashi, Japan
- The University of Shizuoka, Graduate School of Nutritional and Environmental Sciences, Shizuoka, Japan
* Correspondence: Kazuki Mochizuki![]()
Received: December 18, 2017 | Accepted: October 22, 2018 | Published: October 30, 2018
OBM Genetics 2018, Volume 2, Issue 4 doi: 10.21926/obm.genet.1804046
Academic Editors: Stéphane Viville and Marcel Mannens
Special Issue: Epigenetic Mechanisms in Health and Disease
Recommended citation: Mochizuki K, Osaki A, Inamochi Y, oda T. The Induction of Histone H3K4 Methylation on the SI Gene Correlates with SI mRNA Levels in Enterocyte-Like Caco-2 Cells. OBM Genetics 2018;2(4):046; doi:10.21926/obm.genet.1804046.
© 2018 by the authors. This is an open access article distributed under the conditions of the Creative Commons by Attribution License, which permits unrestricted use, distribution, and reproduction in any medium or format, provided the original work is correctly cited.
Abstract
Background: Histone modifications, including acetylation and H3 lysine 4 (K4) methylation, are thought to be associated with transcriptional activation during differentiation. Methods: mRNA and histone modifications around the sucrase-isomaltase (SI) gene in Caco-2 cells (a small intestine cell line) during differentiation were determined by qRT-PCR and chromatin immunoprecipitation, respectively. Results: Mono-, di-, and tri-methylation of histone H3K4 on the SI gene correlated with the induction of SI gene expression, RNA polymerase II binding, and acetylation of histones H3 and H4 close to the transcription initiation site during differentiation in Caco-2 cells. Monomethylation was prominent in the promoter–enhancer region, while dimethylation was observed from the promoter–enhancer region throughout the gene body region. Trimethylation was detected from the transcription initiation site throughout the gene body region. Conclusions: Histone H3K4 methylation was associated with SI gene induction during differentiation in Caco-2 cells. Moreover, the mono-, di-, and tri-methylation of histone H3K4 at different regions of the SI gene may play different roles in SI gene induction.
Keywords
Histone H3K4 methylation; sucrase-isomaltase (SI); differentiation; Caco-2
1. Introduction
Intestinal absorptive cells rapidly differentiate from the crypt to the villus. During differentiation, the cells express many genes related to digestion and absorption. It has been reported that the expression of sucrase-isomaltase (SI), which is an α-glucosidase that digests starch hydrolysates and sucrose to glucose and/or fructose [1], is induced during differentiation from the crypt to the villus [2]. Additionally, many studies have demonstrated that SI expression is induced in the human intestine cell line Caco-2 during differentiation into enterocyte-like cells after contact inhibition [3].
It has been shown in Caco-2 cells that expression of the SI gene is induced by transcription factors such as caudal-related homeobox transcription factor 2 (CDX2), hepatocyte nuclear factor 1 (HNF1), and GATA4/5 [4,5,6,7], which bind to cis-regulatory elements located in the SI promoter region [8]. Indeed, we have recently shown that HNF1 and CDX2 bind to the promoter region of the SI gene, and that this binding correlates with the induction of the SI gene during differentiation from crypt to villus in rats and also upon feeding mice a high-starch diet [9,10]. Furthermore, recent studies suggest that acute gene expression changes, which occur frequently during differentiation, are accompanied by histone modifications such as acetylation, methylation, and phosphorylation [11,12]. Particularly, acetylation of histone H3 lysines (K) 9 and 14 and of histone H4 at K5, K8, K12, and K16 as well as methylation of histone H3K4 are related to transactivation [13,14,15]. Mono-, di-, or tri-methylation of histone H3K4 is associated with the initial transcription activation, because di- or tri-methylation of histone H3K4 induces histone acetylation [16,17]. Histone acetylation connects bromodomain-containing proteins. Subsequently, these proteins recruit transcription complexes to the promoter–enhancer region as well as transcription elongation complexes to the gene body region [18,19,20]. Several studies have found that histone H3K4 trimethylation induces the recruitment of transcription initiation and transcription elongation complexes to target genes by binding proteins containing plant homeodomains (PHD), chromodomains, Tudor domains, or WD40 repeat domains [21,22]. In addition, histone H3K4 methylation induces histone H3K9/14 acetylation [23]. Recently, we have demonstrated in rats and mice that acetylated histones H3 and H4 on the SI gene are correlated with jejunal SI gene induction during the differentiation of intestinal absorptive cells in various settings, i.e., from the crypt to the villus, during postnatal development, and by a high-starch diet or fructose solution [9,10,24]. However, it is not known whether induction of the human SI gene during the differentiation of Caco-2 cells is regulated by histone acetylation. Furthermore, it has not been examined whether induction of the SI gene is associated with methylation of histone H3K4 in intestinal cell lines or in the jejunum in vivo.
In this study, we examined whether the induction of SI mRNA expression during contact inhibition-mediated differentiation of the intestinal cell line Caco-2 is associated with SI gene methylation of histone H3K4 or acetylation of histone H3 at K9/14 and of H4 at K5/8/12/16.
2. Materials and Methods
2.1. Cell Culture
Caco-2 cells, which were purchased from the American Type Culture Collection (Rockville, MD, USA), were seeded in 10-cm collagen-coated culture plates (Iwaki, Tokyo, Japan) at a density of 0.6×104 cells/cm2 with Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal calf serum at 37°C in a humidified atmosphere of 5% CO2. Details of the medium have been described previously [25]. Day 0, which was 5 days after plating, was regarded as the day of 100% confluence.
2.2. RNA Analysis
The total RNA extraction and reverse transcription methods were described previously [25]. cDNAs were amplified by qPCR using a Light Cycler System (Roche Diagnostics, Basel, Switzerland) and SYBR Green I (Takara, Shiga, Japan). The cycle threshold (CT) values of tested genes detected by qRT-PCR such as SI; Solute carrier family 2 member 5 (SLC2A5), which is a facilitated glucose/fructose transporter; alkaline phosphatase, intestinal (ALPI); villin 1 (VIL1); and β-actin (ACTB) were converted into signal intensities by the delta-delta method [26]. The formula used was [2(CT ACTB – CT tested gene)]. The primer sequences for the qRT-PCR are described in Table 1.
2.3. Chromatin Immunoprecipitation (ChIP) Assay
The ChIP method has been described previously [25]. This assay used 1 μg of anti-RNA polymerase II antibody (Pol II) (Covance, MD, USA), anti-acetyl-Histone H3K9/14 antibody (Merck Millipore, Urlington, MA, USA), anti-acetyl-Histone H4K5/8/12/16 antibody (Merck Millipore), anti-monomethyl-Histone H3K4 antibody (Merck Millipore), anti-dimethyl-Histone H3K4 antibody (Merck Millipore), anti-trimethyl-Histone H3K4 antibody (Merck Millipore), or normal rabbit IgG. The CT values of the ChIP signals detected by qPCR were converted to the percentage of the ChIP signal of the input DNA, which was calculated by the delta-delta method [26], using the formula 100×[2(CT IP input – CT sample)]. The primer sequences for the ChIP assays are listed in Table 1.
|
Table 1 The primer sequences for real-time PCR. |
|
|
Target mRNA |
Sequence |
|
SI |
5´-ATTTATACGGCCATCAAACATTCT-3´ |
|
|
5´-TGGAGTAGGCTGGATAAAAATCTC-3´ |
|
SLC2A5 |
5’-TGATCTTCCCGTTCATCCA-3’ |
|
|
5’-GTCTCCGGGACAATCAAGAA-3’ |
|
ALPI |
5´-TGTCTTCTCCTTTGGTGGCTA-3´ |
|
|
5´-ATGGACGTGTAGGCTTTGCT-3´ |
|
VIL |
5´-AGCTTATCAAGCCGTCATCCT-3´ |
|
|
5´-CGTCCCTTGAAGATGGACATA-3´ |
|
ACTB |
5´-CATGAAGTGTGACGTGGACAT-3´ |
|
|
5´-TGATCTCCTTCTGCATCCTGT-3´ |
|
Region around SI gene |
|
|
-10000 |
5´-CATGAGGTCAGGAGTTTGACA-3´ |
|
|
5´-ACCACCGCCTAGCTACTTTTT-3´ |
|
-3200 |
5´-CACAGCGCTAATGCAGAGATA-3´ |
|
|
5´-CAGAGCCTTCTTCCATTCCTT-3´ |
|
-1700 |
5´-GATTCTTTCCAGCTTTCCAC-3´ |
|
|
5´-GGAACATGTAAAGGAGGACT-3´ |
|
-400 |
5´-CCCTGTATCTTGTCTTCGATCT-3´ |
|
|
5´-AAATTACTGATCTGGGCACCTC-3´ |
|
1 |
5´-CTGGTGAGGGTGCAATAAAAC-3´ |
|
|
5´-GACTTGGATAAGGCTGCCAAA-3´ |
|
1100 |
5´-GTAAGGACTCACCATGGCATT-3´ |
|
|
5´-TTGCCTTCATACCTGCTGTTC-3´ |
|
3000 |
5´-CCTTCACACACTTGCAAAGGA-3´ |
|
|
5´-TTCTCTACCTGCCTTTCTCA-3´ |
|
12000 |
5´-AGTATGTGGTGTGGTGGATATG-3´ |
|
|
5´-GGGGATTATCCTGAAAGCAGT-3´ |
|
99800 |
5´-TAACCAACAACTCTGGGCAGT-3´ |
|
|
5´-GGGTGGAGAAAGAAATTGACC-3´ |
|
over5000 |
5´-TCTGCTTTATGCCCCTCACT-3´ |
|
|
5´-GTTATTGGCGGCCTATCCTT-3´ |
|
over10000 |
5´-GGTCAGACTGGCATGTGATTT-3´ |
|
|
5´-CTTGGGGGAAAGGTGAGAATA-3´ |
2.4. Statistical Analysis
Results are expressed as mean ± standard of the error (SEM). Statistical significance was determined by the Kruskal–Wallis multiple range test and the post hoc Dunn’s test.
3. Results
3.1. SI Gene Expression after Confluence in Caco-2 Cells
First, we investigated whether acetylation of histone H3K9/14 and of histone H4K5/8/12/16, as well as methylation of histone H3K4 induces SI gene transcription and RNA Pol II binding to the SI gene during differentiation in Caco-2 cells. As shown in Figure 1, the SI mRNA level was very low at 100% confluence (day 0). However, it increased with time as it was upregulated 14689- and 75029-fold at days 5 and 10, respectively, compared with day 0. Furthermore, other intestinal differentiation markers showed higher expression on days 5 and 10 than on day 0, including SLC2A5, which is a fructose transporter, ALPI, which is related to digestion of phosphate compounds, and VIL1, which is involved in villus construction.
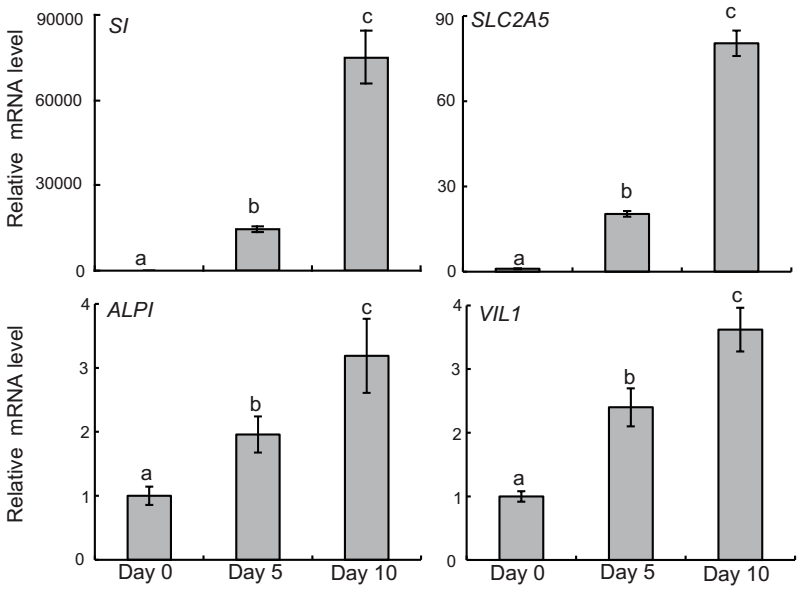
Figure 1 Intestinal gene expression in Caco-2 cells during differentiation. Samples were collected on days 0, 5, and 10 after confluency for qRT-PCR. Results of qRT-PCR were normalized to ACTB mRNA. Values with different superscript letters differ significantly (P < 0.05).
3.2. Association of Pol II, Acetylation of Histone H3K9/14 and of Histone H4K5/8/12/16, and Methylation of Histone H3K4 with SI Gene Induction during Caco-2 Differentiation
We performed ChIP assays using antibodies for acetylated histone H3K9/14, acetylated histone H4K5/8/12/16, Pol II, and mono/di/trimethylated histone H3K4 to examine whether these histone modifications correlate with the induction of SI mRNA and with Pol II binding to the SI gene during differentiation in Caco-2 cells. The primer positions for the ChIP assays are described in Figure 2A and Figure 3A. The ChIP signals of normal rabbit IgG were under 0.04% per input. The signals for acetylation of histone H3K9/14 at 1100 bp was notably enhanced during differentiation (Figure 2B). The signals of acetylated histone H4K5/8/12/16 were pronouncedly enhanced after confluence between days 0 and 10, particularly in the promoter–enhancer region (–1700 bp to –400 bp) and at 1 bp to 1100 bp (Figure 2C). The bindings of Pol II were markedly enhanced after confluence, particularly in regions close to the transcription initiation site; significant induction was observed between days 0 and 10 between –400 bp and 1 bp (Figure 2D).
Regarding the methylation of histone H3K4, the signals of monomethylated histone H3K4 were considerably enhanced after confluence, particularly in the promoter–enhancer region (–1700 bp and –400 bp); significant induction was observed between days 0 and 10 at –1700 bp, –400 bp, 1100 bp, and 3000 bp (Figure 3B). The signals of dimethylated histone H3K4 were significantly higher on day 10 than on day 0 at the promoter–enhancer region and throughout the gene body region (from –3200 bp to 3000 bp; Figure 3C). Finally, the signals of trimethylated histone H3K4 were markedly enhanced after confluence, particularly in regions close to the transcription initiation site (–400 bp and 1 bp), but also in the rest of the gene body region (1100 bp and 3000 bp), peaking at 1100 bp (Figure 3D).

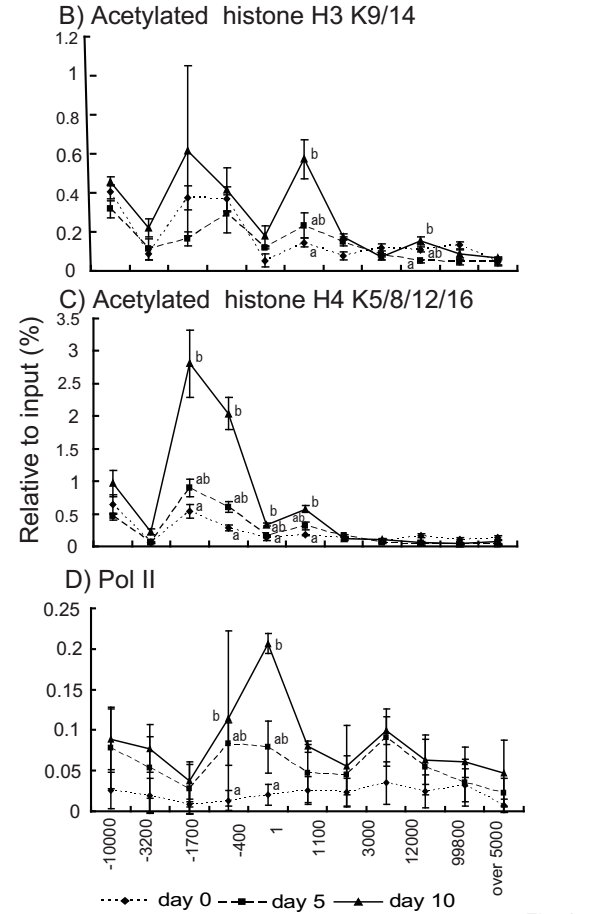
Figure 2 Acetylated histone H3K9/14, acetylated histone H4K5/8/12/16, and binding of Pol II to the SI gene in Caco-2 cells during differentiation. (A) Positions of primers on the SI gene. (B–D) Chromatin immunoprecipitation (ChIP) assays for acetylated histone H3K9/14 (B), acetylated histone H4K5/8/12/16 (C), and Pol II (D). The samples were collected for ChIP assays on days 0, 5, and 10 after confluence. ChIP signals were detected by qPCR and were normalized to the input signals. The abscissae denote the region on the SI gene relative to the transcription initiation site. Values with different superscript letters differ significantly (P < 0.05).
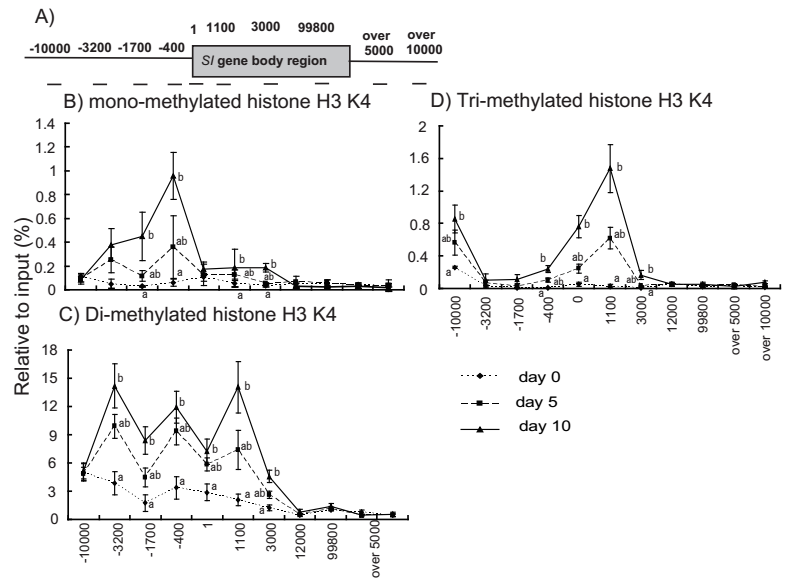
Figure 3 Binding of methylated histone H3K4 to the SI gene in Caco-2 cells during differentiation. (A) Positions of primers on the SI gene. (B–D) Chromatin immunoprecipitation (ChIP) assays for monomethylated histone H3K4 (B), dimethylated histone H3K4 (C), and trimethylated histone H3K4 (D). The samples were collected for ChIP assays on days 0, 5, and 10 after confluence. ChIP signals were detected by qPCR and were normalized to the input signals. The abscissae denote the region on the SI gene relative to the transcription initiation site. Values with different superscript letters differ significantly (P < 0.05).
4. Discussion
In this study, we showed that SI gene expression was upregulated during differentiation induced by contact inhibition in Caco-2 cells. It has been reported that the SI protein level was not detected before confluence and increased exponentially after confluence in Caco-2 cells [27]. This report is consistent with the SI mRNA levels in Caco-2 cells observed in the current study. Here, we demonstrated that acetylation of histone H3K9/14 within the gene body region and of histone H4K5/8/12/16 in the promoter–enhancer region as well as Pol II binding to the region close to the transcription initiation site of the SI gene were all progressively increased by differentiation. Because the hyperacetylation of histone H3 correlates with the euchromatin region in the genome [12,14,28], our results in this current study suggest that histone acetylation is concerned with SI gene expression during the differentiation of Caco-2 cells and that this acetylation is associated with the induction of the gene in humans. Furthermore, we showed that histone H3 was highly acetylated in the gene body region, although histone H4 was highly acetylated in the promoter–enhancer region. Our previous studies on histone acetylation of the SI gene during postnatal development of rats or while feeding mice a high-starch or fructose diet showed similar results [10,24,29]. Histone acetylation in the gene body region enhances the transcription elongation by recruiting transcription elongation complexes, whereas histone acetylation in the promoter–enhancer region enhances transcription initiation by recruiting mRNA transcription initiation complexes [18,19,20]. Therefore, it is very likely that acetylation of histone H3 and H4 is involved in the regulation of the transcription elongation reaction and of the transcription initiation reaction, respectively.
In this study, we found that trimethylation of histone H3K4 was induced during differentiation in the region close to the transcription initiation site and the gene body region of the SI gene. Previous studies have demonstrated that histone H3K4 was highly trimethylated in the region close to and just after the transcription initiation site, which has a role in recruiting Pol II and enhancing transcription elongation [30,31]. Our results are in accordance with these studies, because Pol II binding and histone H3K4 trimethylation were induced during differentiation, and correlated with SI gene induction. Furthermore, we revealed that dimethylation from the promoter–enhancer region throughout the gene body region and monomethylation in the promoter–enhancer region were increased during differentiation. It has been reported that both di- and tri-methylation are found predominately on active loci, although trimethylation occurs concomitantly with active transcription, while dimethylation can be found on poised, inactive genes [23,32]. It should be noted that we detected high levels of dimethylation in the promoter–enhancer and gene body regions of the SI gene, even on day 0 when SI expression was not observed, whereas trimethylation was closely associated with the induction of the SI gene. The promoter–enhancer region is important for the initiation of chromatin remodeling because CpG islands are located in the promoter region, and many transcription factors and transcription complexes are recruited to the promoter–enhancer region for the initiation of transcription. We hypothesized that mono- and di-methylation of histone H3K4 may function in chromatin remodeling when preparing to change from heterochromatin to euchromatin as well as in recruiting complexes that are required prior to the initial stages of mRNA transcription and elongation.
It should be noted that the mRNA level of the SI gene was pronouncedly higher at day 5 after confluence than at day 0, although the differences of histone modifications and Pol II binding around the SI gene between days 0 and 5 were smaller. The difference may be due to the sensitivity of the ChIP assay. Another possibility is that induction of the SI gene was regulated by other factors including transcriptional factors; transcription of SI mRNA in Caco-2 cells is reportedly induced by several transcription factors such as CDX2, HNF1, and GATA4/5 in the SI promoter region [4,5,6,7]. It should be examined in further studies whether bindings of transcription factors around the SI gene are induced during differentiation of the Caco-2 cells.
In conclusion, we found that histone H3K4 methylation was associated with SI gene induction, Pol II binding, and histone acetylation on the SI gene during differentiation of the human enterocyte-like cell line, Caco-2. We demonstrated for the first time that the modification of histone H3K4 from the promoter–enhancer region to the gene body region of an intestinal gene gradually shifted from mono- to tri-methylation.
Acknowledgments
We thank Michal Bell, PhD, from Edanz Group (www.edanzediting.com/ac) for editing the English text of a draft of this manuscript.
Author Contributions
KM did experiments and wrote the manuscript, AO and YI did experiments, TG helped the writing the manuscript.
Funding
This work was supported by Grants-in-Aid for Young Scientists (22680054) and for Scientific Research (26282023) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), and by a grant from The Uehara Memorial Foundation.
Competing Interests
The authors have declared that no competing interests exist.
References
- Semenza G. Anchoring and biosynthesis of stalked brush border membrane proteins: glycosidases and peptidases of enterocytes and renal tubuli. Annu Rev Cell Biol. 1986; 2: 255-313. [CrossRef]
- Goda T, Yasutake H, Tanaka T, Takase S. Lactase-phlorizin hydrolase and sucrase-isomaltase genes are expressed differently along the villus-crypt axis of rat jejunum. J Nutr. 1999; 129: 1107-1113. [CrossRef]
- Van Beers EH, Al RH, Rings EH, Einerhand AW, Dekker J, Buller HA. Lactase and sucrase-isomaltase gene expression during Caco-2 cell differentiation. Biochem J. 1995; 308: 769-775. [CrossRef]
- Olsen WA, Lloyd M, Korsmo H, He YZ. Regulation of sucrase and lactase in Caco-2 cells: relationship to nuclear factors SIF-1 and NF-LPH-1. Am J Physiol. 1996; 271: G707-713.
- Rodolosse A, Carriere V, Rousset M, Lacasa M. Two HNF-1 binding sites govern the glucose repression of the human sucrase-isomaltase promoter. Biochem J. 1998; 336: 115-123. [CrossRef]
- Boudreau F, Rings EH, van Wering HM, Kim RK, Swain GP, Krasinski SD, et al. Hepatocyte nuclear factor-1α, GATA-4, and caudal related homeodomain protein Cdx2 interact functionally to modulate intestinal gene transcription. Implication for the developmental regulation of the sucrase-isomaltase gene. J Biol Chem. 2002; 277: 31909-31917. [CrossRef]
- Krasinski SD, Van Wering HM, Tannemaat MR, Grand RJ. Differential activation of intestinal gene promoters: functional interactions between GATA-5 and HNF-1α. Am J Physiol Gastrointest Liver Physiol. 2001; 281: G69-84. [CrossRef]
- Traber PG, Wu GD, Wang W. Novel DNA-binding proteins regulate intestine-specific transcription of the sucrase-isomaltase gene. Mol Cell Biol. 1992; 12: 3614-3627. [CrossRef]
- Suzuki T, Mochizuki K, Goda T. Histone H3 modifications and Cdx-2 binding to the sucrase-isomaltase (SI) gene is involved in induction of the gene in the transition from the crypt to villus in the small intestine of rats. Biochem Biophys Res Commun. 2008; 369: 788-793. [CrossRef]
- Honma K, Mochizuki K, Goda T. Carbohydrate/fat ratio in the diet alters histone acetylation on the sucrase-isomaltase gene and its expression in mouse small intestine. Biochem Biophys Res Commun. 2007; 357: 1124-1129. [CrossRef]
- Rice JC, Briggs SD, Ueberheide B, Barber CM, Shabanowitz J, Hunt DF, et al. Histone methyltransferases direct different degrees of methylation to define distinct chromatin domains. Mol Cell. 2003; 12: 1591-1598. [CrossRef]
- Schubeler D, MacAlpine DM, Scalzo D, Wirbelauer C, Kooperberg C, van Leeuwen F, et al. The histone modification pattern of active genes revealed through genome-wide chromatin analysis of a higher eukaryote. Genes Dev. 2004; 18: 1263-1271. [CrossRef]
- Yan C, Boyd DD. Histone H3 acetylation and H3 K4 methylation define distinct chromatin regions permissive for transgene expression. Mol Cell Biol. 2006; 26: 6357-6371. [CrossRef]
- Roh TY, Cuddapah S, Zhao K. Active chromatin domains are defined by acetylation islands revealed by genome-wide mapping. Genes Dev. 2005; 19: 542-552. [CrossRef]
- Lin W, Dent SY. Functions of histone-modifying enzymes in development. Curr Opin Genet Dev. 2006; 16: 137-142. [CrossRef]
- Wang H, Cao R, Xia L, Erdjument-Bromage H, Borchers C, Tempst P, et al. Purification and functional characterization of a histone H3-lysine 4-specific methyltransferase. Mol Cell. 2001; 8: 1207-1217. [CrossRef]
- Musri MM, Corominola H, Casamitjana R, Gomis R, Parrizas M. Histone H3 lysine 4 dimethylation signals the transcriptional competence of the adiponectin promoter in preadipocytes. J Biol Chem. 2006; 281: 17180-17188. [CrossRef]
- Kanno T, Kanno Y, Siegel RM, Jang MK, Lenardo MJ, Ozato K. Selective recognition of acetylated histones by bromodomain proteins visualized in living cells. Mol Cell. 2004; 13 :33-43. [CrossRef]
- Jang MK, Mochizuki K, Zhou M, Jeong HS, Brady JN, Ozato K. The bromodomain protein Brd4 is a positive regulatory component of P-TEFb and stimulates RNA polymerase II-dependent transcription. Mol Cell. 2005; 19: 523-534. [CrossRef]
- Yang XJ. Lysine acetylation and the bromodomain: a new partnership for signaling. Bioessays. 2004; 26: 1076-1087. [CrossRef]
- Sims RJ, 3rd, Reinberg D. Histone H3 Lys 4 methylation: caught in a bind? Genes Dev. 2006; 20: 2779-2786. [CrossRef]
- Wysocka J, Swigut T, Xiao H, Milne TA, Kwon SY, Landry J, et al. A PHD finger of NURF couples histone H3 lysine 4 trimethylation with chromatin remodelling. Nature. 2006; 442: 86-90. [CrossRef]
- Bernstein BE, Kamal M, Lindblad-Toh K, Bekiranov S, Bailey DK, Huebert DJ, et al. Genomic maps and comparative analysis of histone modifications in human and mouse. Cell. 2005; 120: 169-181. [CrossRef]
- Honma N, Mochizuki K, Goda T. Acute induction of histone acetylation on the jejunal sucrase–isomaltase gene by dietary fructose. Br J Nutr. 2008; 100: 698-702. [CrossRef]
- Inamochi Y, Mochizuki K, Osaki A, Ishii T, Nakayama T, Goda T. Histone H3 methylation at lysine 4 on the SLC2A5 gene in intestinal Caco-2 cells is involved in SLC2A5 expression. Biochem Biophys Res Commun. 2010; 392: 16-21. [CrossRef]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001; 25: 402-408. [CrossRef]
- Lemieux E, Boucher MJ, Mongrain S, Boudreau F, Asselin C, Rivard N. Constitutive activation of the MEK/ERK pathway inhibits intestinal epithelial cell differentiation. Am J Physiol Gastrointest Liver Physiol. 2011; 301: 719-30. [CrossRef]
- Roh TY, Ngau WC, Cui K, Landsman D, Zhao K. High-resolution genome-wide mapping of histone modifications. Nat Biotechnol. 2004; 22: 1013-1016. [CrossRef]
- Yorita S, Mochizuki K, Goda T. Induction of histone acetylation on the sucrase-isomaltase gene in the postnatal rat jejunum. Biosci Biotechnol Biochem. 2009; 73: 933-935. [CrossRef]
- Ng HH, Robert F, Young RA, Struhl K. Targeted recruitment of Set1 histone methylase by elongating Pol II provides a localized mark and memory of recent transcriptional activity. Mol Cell. 2003; 11: 709-719. [CrossRef]
- Santos-Rosa H, Schneider R, Bannister AJ, Sherriff J, Bernstein BE, Emre NC, et al. Active genes are tri-methylated at K4 of histone H3. Nature. 2002; 419: 407-411. [CrossRef]
- Schneider R, Bannister AJ, Myers FA, Thorne AW, Crane-Robinson C, Kouzarides T. Histone H3 lysine 4 methylation patterns in higher eukaryotic genes. Nat Cell Biol. 2004; 6: 73-77. [CrossRef]

